We have obtained x-ray scattering data at high instrumental resolution
for the ![]() (fluid,
chain melted) phase of DPPC at 50
(fluid,
chain melted) phase of DPPC at 50![]() C
for samples exhibiting a wide variety of D-spacings, as summarized in Table
I. Smaller D-spacings occurred when higher osmotic pressure was applied
by increasing the concentration of PVP. The maximum number
C
for samples exhibiting a wide variety of D-spacings, as summarized in Table
I. Smaller D-spacings occurred when higher osmotic pressure was applied
by increasing the concentration of PVP. The maximum number ![]() of observable small angle lamellar scattering peaks increased as D decreased.
Incidentally, we have consistently found that the D-spacing of fully hydrated
DPPC (with 0% PVP) varies noticeably from sample to sample. The first two
samples in Table I give the maximum range that we have observed, although
it may be noted that Shipley's group (Janiak et al, 1976) has observed
D-spacings as low as 60.0 Ĺ.
of observable small angle lamellar scattering peaks increased as D decreased.
Incidentally, we have consistently found that the D-spacing of fully hydrated
DPPC (with 0% PVP) varies noticeably from sample to sample. The first two
samples in Table I give the maximum range that we have observed, although
it may be noted that Shipley's group (Janiak et al, 1976) has observed
D-spacings as low as 60.0 Ĺ.
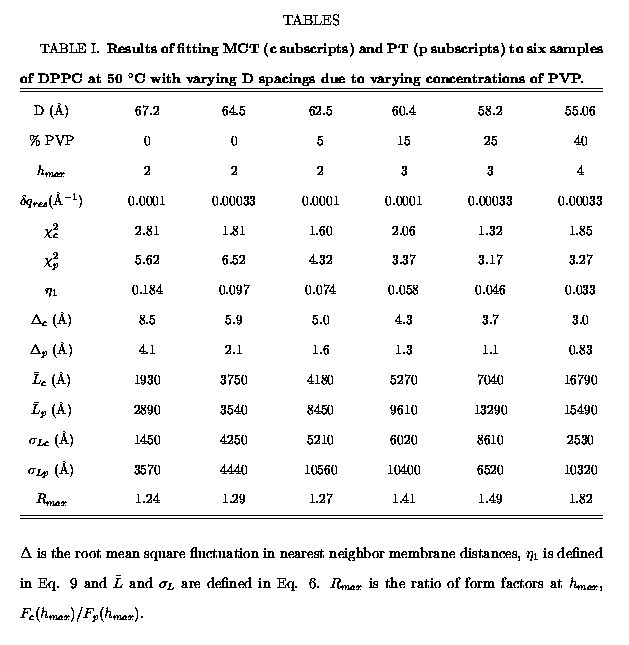
Figure 5 shows the scattering data for
the DPPC sample with 0% PVP and D = 64.5 Ĺ. Although these data were taken
with the wider longitudinal resolution using the dispersive detector set-up,
the peak shapes are well resolved. The PT fit to the first order peak appears
visually to be nearly as good as the MCT fit, but the MCT fit to the second
order peak is clearly superior. As can be seen in Table I, the ratio of
the ![]() for the
two theories is 3.6, indicating that MCT is far superior to PT.
for the
two theories is 3.6, indicating that MCT is far superior to PT.
Figure 6 shows the scattering for the
other 0% PVP sample with D = 67.2 Ĺ. These data were taken with our highest
resolution. The scattering for ![]() Ĺ
Ĺ![]() appears to
be anomalously high and erratic; these data points are not fit well by
either theory and this accounts for the relatively large values of
appears to
be anomalously high and erratic; these data points are not fit well by
either theory and this accounts for the relatively large values of ![]() .
Nevertheless, the ratio of
.
Nevertheless, the ratio of ![]() for the two theories strongly supports the conclusion that MCT is better
than PT.
for the two theories strongly supports the conclusion that MCT is better
than PT.
Figure 7 shows scattering data for
the 25% PVP sample with D = 58.2 Ĺ. Again, although there is little difference
in the fits to the first order peak, MCT clearly fits both higher order
peaks better than PT. Data (not shown) for the other three samples confirms
this general conclusion. We have also analysed much additional data from
DPPC using only MCT with similar results that will be reported in a subsequent
publication. MCT fits to less extensive data from DMPC have previously
been shown by Zhang et al. (1995); these latter fits gave smaller
![]() values near
1.4 by relaxing the constraint in Eq. 10.
values near
1.4 by relaxing the constraint in Eq. 10.
The best fits to the data yield the values of the parameters reported
in Table I. As expected, the values of ![]() ,
,
![]() and
and ![]() decrease monotonically with decreasing D as fluctuations in the multilamellar
vesicles are suppressed by increased osmotic pressure. Table I shows that
the root mean square fluctuations in nearest neighbor distances, which
are given by
decrease monotonically with decreasing D as fluctuations in the multilamellar
vesicles are suppressed by increased osmotic pressure. Table I shows that
the root mean square fluctuations in nearest neighbor distances, which
are given by ![]() for MCT (from Eq. 11) and by
for MCT (from Eq. 11) and by ![]() from the PT fit, are predicted to be larger by MCT than by PT, by a factor
that varies from about 2 to nearly 4 as D decreases. The probable uncertainties
in the parameters
from the PT fit, are predicted to be larger by MCT than by PT, by a factor
that varies from about 2 to nearly 4 as D decreases. The probable uncertainties
in the parameters ![]() and
and ![]() are quite
small, of order 1%. The mean domain sizes
are quite
small, of order 1%. The mean domain sizes ![]() are quite large for both theories, corresponding to average number N of
bilayers per domain from 30 to 300. Since the values of
are quite large for both theories, corresponding to average number N of
bilayers per domain from 30 to 300. Since the values of ![]() are comparable to the values of
are comparable to the values of ![]() for both theories, the domain size distribution is quite broad.
for both theories, the domain size distribution is quite broad.
The final row in Table I indicates how the form factors that arise from
fits to the data using MCT and PT differ. Specifically, the form factors
F(1) for the first order peaks are normalized to unity for the fits from
both theories. The fits to the peak shapes yield S(q) shapes with amplitudes
that give F(h) according to Eq. 4. Finally,
for the highest order peak, ![]() ,
the ratio,
,
the ratio, ![]() ,
of the MCT form factor to the PT form factor is reported in Table I.
,
of the MCT form factor to the PT form factor is reported in Table I.
Finally, it may be of interest to see how our modifications to the Caille
theory affect fits to the data compared to the unmodified Caille theory
that was used earlier by Roux and Safinya (1988). Previous authors did
not report ![]() values and it is difficult in any case to compare different experiments
directly. However, we have performed fits to some of our data using the
unmodified Caille theory, as shown in Fig. 8. The
values and it is difficult in any case to compare different experiments
directly. However, we have performed fits to some of our data using the
unmodified Caille theory, as shown in Fig. 8. The ![]() for the unmodified Caille theory is much larger than the
for the unmodified Caille theory is much larger than the ![]() for the modified theory. The unmodified theory does a good job fitting
the tails; the primary reason for the larger
for the modified theory. The unmodified theory does a good job fitting
the tails; the primary reason for the larger ![]() is the difficulty in fitting the sharp first order peak and this is primarily
due to the classical finite size factor which is an artificial approximation
(Zhang et al., 1994). It is interesting to compare the unmodified Caille
theory to PT which fits the h=1 peak better (Fig. 5) but has trouble with
the h=2 peak for which the tails are larger.
is the difficulty in fitting the sharp first order peak and this is primarily
due to the classical finite size factor which is an artificial approximation
(Zhang et al., 1994). It is interesting to compare the unmodified Caille
theory to PT which fits the h=1 peak better (Fig. 5) but has trouble with
the h=2 peak for which the tails are larger.